CASA-CNP
The Tools
From the CASA-CNP model, only the carbon-cycle equations are used in BIOS2, with the nitrogen and phosphorous cycles disabled. For BIOS2, additional CASA-CNP modifications, made to improve model performance against observations, include using static allocation coefficients (rather than allocation coefficients dependent upon phenology, temperature, and soil moisture), and holding the ratio of NPP to GPP constant in time, instead of using the default growth respiration/maintenance respiration paradigm which is known to be problematic.
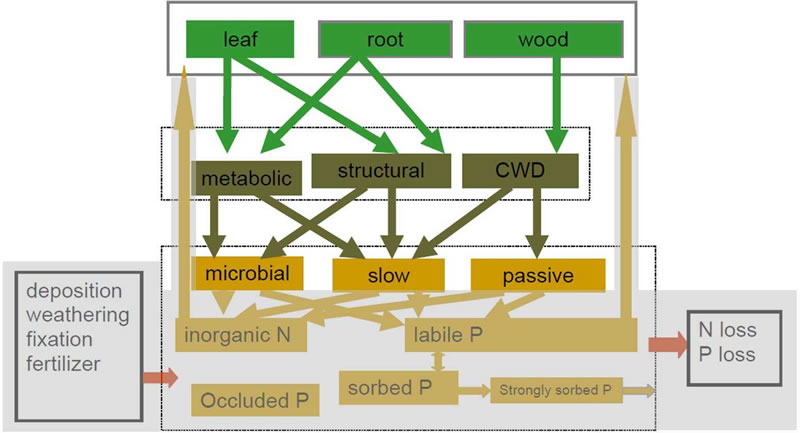
The carbon cycle of CASA-CNP (Wang et al. 2010) is based on the CASA’ model (Fung et al., 2005). Wang et al. reduced the number of carbon pools by combining surface litter with soil litter, and surface microbial biomass with soil microbial biomass. This gives three discrete pools in the litter: structural, metabolic and coarse woody debris pools and three organic pools in the soil: microbial biomass, slow and passive pools. The fluxes between different pools are modelled as in CASA’.
Transfer coefficients from plant pool to litter pool and from litter pool to soil pool are calculated as in the CASA’ model (Fung et al., 2005). Turnover rates of litter carbon or soil carbon are a function of substrate quality (lignin:N ratio), soil temperature, moisture and soil texture (Randerson et al., 1997). The turnover rate of leaves is calculated as a function of leaf age (Arora and Boer, 2005), and the turnover rates of woody tissue or fine roots are constant for each biome, but vary with biome type.